Video & notes: Kangaroos, culling and Canberra - presentation to AJP event 5 April 2016
 Video and text of Sheila Newman's speech at the Animal Justice Party's event, "Policy basis for Kangaroo treatment in the ACT," 5 April 2016: Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover. Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes. Migration has probably been mistaken for fertility, rendering ACT roo counts unreliable and invalid. The ACT needs to stop culling and widen its research base to consider various genetically based algorithms that naturally restrain fertility opportunities in kangaroos.
Video and text of Sheila Newman's speech at the Animal Justice Party's event, "Policy basis for Kangaroo treatment in the ACT," 5 April 2016: Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover. Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes. Migration has probably been mistaken for fertility, rendering ACT roo counts unreliable and invalid. The ACT needs to stop culling and widen its research base to consider various genetically based algorithms that naturally restrain fertility opportunities in kangaroos.
Examples include separate gender pathways, with 'sexual segregation' where male and female populations live apart. It is likely that the stable presence of mature dominant males and females in family and mob organisation inhibits sexual maturity and activity as has been shown in studies of other species, such as macaques and superb fairy wrens (the latter cooperative breeders). In humans, girls brought up with step-fathers who came late to the family were more likely to mature sexually earlier due to absence of Westermarck Effect.)
Planned wildlife corridors need to be made safe and long-term viable to cope with people, car and kangaroo population movements.
Canberra is pursuing a policy of rapid population growth, mostly through invited economic immigration.
Canberra's population problem

In June 2016 ACT - South West Australian Capital Territory was the fastest growing area in Australia and grew by 127.3%. (ABS http://www.abs.gov.au/ausstats/[email protected]/mf/3218.0)
Canberra’s population could increase to 904,000 by 2061 according to new projections released by the Australian Bureau of Statistics. It's not inevitable, but the government would like you to believe it is.
Predicting a population growth of at least 98 per cent within 50 years, ACT population projections for 2061 suggest that the Australian Capital Territory population could exceed Tasmania's population by 2038.
But some think that the ACT's biggest problem is its kangaroo population
It's not the new suburbs, the new roads, the new airport, the additional schools, hospitals, houses, and all the new cars that threaten Canberra's grasslands: it's the eastern grey kangaroos.
It seems that it is better to have cattle in Canberra's nature parks than kangaroos.
Ecological cattle grazing is now being trialed. Cattle can be more easily moved than kangaroos. (Fletcher, Senior Ecologist, communication to P. Machin.)

Although, Fletcher had previously described the devastation cattle made to grass cover in no uncertain terms: "Fletcher Phd: p.37. “70 pregnant cows and four bulls grazed for ten weeks at Tidbinbilla after a bushfire in January 2003 (Section 3.5.1). Prior to their arrival, there had been an atypical abundance of pasture due to the death of almost half of the Tidbinbilla kangaroos in the bushfire, but by the time the cattle were removed, the Tidbinbilla pasture had been reduced to the lowest herbage mass recorded on any site during the study.”
"

ACT Kangaroo Management Policy works on a model that all creatures maximize their population growth and that Canberra's roos are riding an expansive curve which can only be capped by massive frequent culls. A stated fear is that they will otherwise graze and drastically modify biodiversity of Canberra's grasslands. Another is that roos need periodically to be shot so as to save them from starving to death.
Why culling is better than harvesting (ACT Senior Ecologist, Don Fletcher)
"[…] the model indicates that commercial harvesting (currently under trial in the region, at the maximum level allowed, results in a sustainable harvest of kangaroos, but does not increase the herbage mass, and only slightly reduces the frequency of crashes when herbage mass falls to low levels. (To demonstrate this with an ecological experiment would require an extremely large investment of research effort.)
However, an alternative 'national park damage mitigation' formula, which holds kangaroo density to about 1 ha -1 , is predicted to increase herbage mass considerably and to reduce the frequency of crashes in herbage mass, but these effects would be achieved at the cost of having to shoot large numbers of kangaroos." (Fletcher Phd: Population dynamics of Eastern Grey Kangaroos in Temperate Grasslands, 2006, p. vi.)
The model and the reality
The ACT Roo Management model is one of high fertility sedentary populations that rarely migrate, grazing grasslands down to the subsoil. But, in his 2006 thesis, p. 237, Senior ACT Kangaroo management ecologist, Donald Fletcher, tested this model and found, to his surprise, that,
"The study did not provide evidence that high densities of kangaroos reduce groundcover to the levels where erosion can accelerate.
Unmanaged kangaroo populations did not necessarily result in low levels of ground cover. Groundcover had a positive but not significant relationship to kangaroo density, with the highest cover at the wettest site where kangaroo density was highest. Weather has an important influence on groundcover."
"The results from the study as a whole indicate that unmanaged kangaroo populations did not necessarily result in unacceptably low levels of ground cover." (Fletcher Phd: p.231.
The Kangaroo Migration factor
Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes.
Migration has probably been mistaken for fertility, rendering ACT kangaroo counts unreliable and probably invalid.
DNA studies 2003 show migration a strong factor
Zenger et al (2003)[1] found that mitrochondrial DNA samples indicated about 22.61 individuals per generation migrated with a range of 8.17-59.30. In female immigrants the range was 2.73 with a range of 0.60-12.16. Although females demonstrate smaller migration rates compared to the sexes combined, the values are still comparatively high. Analysis across NSW showed populations separated by up to about 230km had equivalent numbers of close relatives when compared to populations only about 20km apart.
This contradicted field study opinion that migration was low in eastern grey kangaroos, and especially low in females in the ACT. Tidbinbilla (a Canberra nature park studied by Fletcher) featured in the Zenger et al study.
Zenger MtDNA findings contrary to Migration views in Fletcher thesis
"Throughout their lives eastern grey kangaroos are relatively sedentary (Johnson 1989) compared to red kangaroos (Priddel 1987). A partly concurrent study of eastern grey kangaroo habitat use and movements on the Googong and Tidbinbilla sites found the eastern grey kangaroos on these sites were sedentary in all seasons (minimum convex polygon mean size 0.43 km 2 ± 0.06 SE and 0.61 km 2 ± 0.08 respectively; Viggers and Hearn 2005). Kangaroos were not radio tracked at Gudgenby but my observations suggest there is no more movement of eastern grey kangaroos on and off the site there than at Tidbinbilla. Thus it is likely there was little net movement of kangaroos on and off the study sites." (Fletcher, page v.)
Paradoxical impact of Culls, kills and Harvests
Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover.
Harvested stock grow smaller, breed earlier, faster
"Smaller, earlier breeding genetic stock tend to escape harvesting". See, e.g. J.J. Poosa, A. Brannstrom, U. Dieckmann, “Harvest-induced maturation evolution under different life-history trade-offs and harvesting regimes.” (See note for more literature on this.)[2]
Fletcher, on estimates of biomass consumption per roo allows for large variations in harvested populations vs wild populations.
"How big are eastern grey kangaroos?
The mean live weight of eastern grey kangaroos taken from the unshot population at Tidbinbilla was 29 kg – smaller than the 35 kg mean live weight assumed in the Kinchega kangaroo study (Caughley et al. 1987). Based on the size relationship between shot and unshot populations of kangaroos in South Australia and Queensland (Grigg 2000), the mean size of eastern grey kangaroos in equivalent shot populations was predicted to be 17 kg live weight. The minimum dressed size accepted by operators of commercial chillers is 17 kg, implying that many of the kangaroos in shot populations (on rural properties) in the ACT region are too small to attract commercial shooters."(Fletcher, p.242.)
Culling has a similar effect.
Earlier maturation would contribute to higher population growth rates. What role does harvesting, culling and farm mitigation killing play in accelerating breeding rates?
“Smaller, eat less, more numerous, more fecund", reproduce earlier
“The management implications arising from this study are numerous and a full account would require a separate report. As one example, kangaroos in these temperate grasslands are on average smaller, eat less, are more numerous, and are more fecund, than would be predicted
from other studies (e.g. Caughley et al. 1987). Thus the benefit of shooting each kangaroo, in terms of grass production, is less, or, in other words, more kangaroos have to be shot to achieve a certain level of impact reduction, and the population will recover more quickly, than would have been predicted prior to this study.” (Fletcher, p245.)
Kangaroos shot in Tidbinbilla and low weight in shot populations
“The mean live weight of eastern grey kangaroos in high density populations can be estimated from the weights of a sample of 332 kangaroos shot at Tidbinbilla in June 1997 (Graeme Coulson, personal communication, 2003) to be 29 kg. (That is an adjustment of the actual mean liveweight of the shot sample, 26.4 kg, to allow for seasonal effects, as explained in Discussion. Kangaroos in shot populations, such as on grazing properties, are likely to be smaller due to selective harvesting, also explained in Discussion).”(Fletcher p.242.)
Wider Research Base needed
In my view, the ACT needs to stop culling and widen its research base to consider encouraging various genetically based behaviours that naturally restrain fertility opportunities in roos.
Known examples include
incest avoidance, which limits breeding unless animals can disperse to their own territory. [3]
Sexual segregation and gender pathways, where male and female populations live apart.
Incest avoidance as a spatial limiter of breeding opportunity
Many examples of suppressed maturity or breeding in both males and females close by related adults in many species. (Sheila Newman, Demography Territory Law: Rules of animal and human populations, Countershock Press, 2013, chapter 3.) (Paperback edition and Kindle edition.)
In kangaroos male sexual dominance and monopolisation of females is a very obvious trait. (The effect of dominant close females on female maturation is less known and should be investigated as it has been in other species).
Where large males and females are removed from mobs, these limiting population effects are also removed. What happens then?
Kinship rules and incest avoidance
The following diagram is of human kinship rules, however similar patterns of incest avoidance occur in other species, and in kangaroos. The diagram for humans is split into family and in-laws and sets out some typical rules for incest avoidance in low fertility environments - central Australia and mountainous South Korea. The rules for inlaws are reproduced back to front to demonstrate a mirror-like effect. The person in the top left corner, 'You" may not conceive/marry any of the people in the black squares. That leaves only eight possible mates - as long as they are not already married. Imagine how hard it would be to find a wife or husband under these circumstances in a sparsely populated society of small clans that only travelled on foot, without cars, planes or boats. In a more fertile environment, the rules of incest avoidance are usually much less strict, as in Leviticus, where people may marry their first cousins - giving much greater fertility opportunities, even without the benefit of modern transport. For more about this and how it affects human economies see: "Overpopulation: Endogamy,Exogamy and fertility opportunity theory"

The following two diagrams are from Zenger et al (2003).
They show the regions from which their eastern grey kangaroo DNA samples were taken, and they give a 'family tree' of roo DNA diversity, which shows greatly decreased diversity in north NSW and in Queensland. The authors could find no explanation for this.

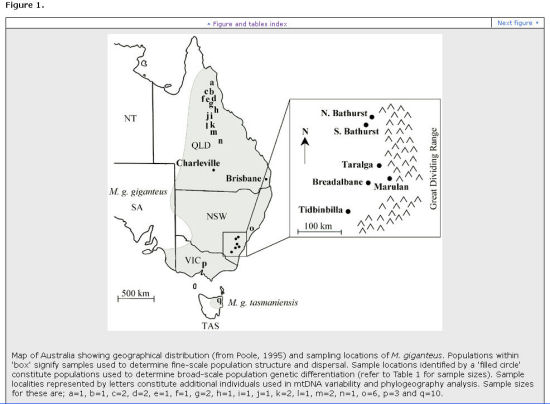
Possible explanation for decreased Mt DNA in Queensland & Northern NSW
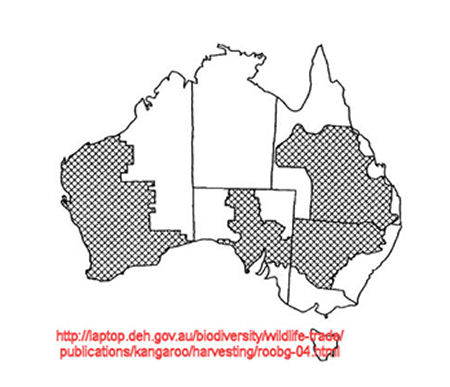
Harvesting has gone on for a long time in these regions. We know it is associated with marked size decrease. It seems likely that it is also associated with earlier sexual maturity. Consider the possibility that, as well as size decrease and earlier sexual maturation in harvested populations, the decrease in genetic diversity present in those populations may be due to inbreeding resulting from loss of family structure and associated incest avoidance, with decreased migration as small early maturing roos settle for their siblings. There seems little will to investigate this. Although there is some literature, it is very limited. (See note [2].)
Sexual Segregation/Gender pathways
Review of Scientific Literature Relevant to the Commercial Harvest Management of Kangaroos http://www.environment.nsw.gov.au/resources/nature/110641Kangaroolitreview.pdf
“Sexual segregation is a phenomenon seen in many species, with segregation occurring along behavioural or ecological dimensions. Sexual segregation in western grey and red kangaroos in semi - arid Victoria has been the subject of intensive investigations since the last review.
[…] MacFarlane and Coulson (2005) investigated the effects of mating activity, group […] composition, spatial distribution and habitat selection on sexual segregation in western grey and red kangaroos. The synchrony and timing of mating activity was seen to influence the magnitude and timing of social segregation in these species, with mixed sex groups predominating during the breeding season. …
… Spatial segregation and habitat segregation were also seen. Although the magnitude of these types of segregation were weaker, they were both still significantly influenced by synchrony and timing of breeding.
Coulson et al. (2006) discussed sexual segregation at three levels (habitat, social and dietary) and confirmed that both size and sex influence segregation.
MacFarlane and Coulson (2009) showed that the need for males to maintain contact with other males (perhaps to develop important fighting skills, evaluate rivals and establish a dominance hierarchy) might also promote sexual segregation.
Similarly Nave (2002) reported evidence of sexual segregation in eastern grey kangaroos in Victoria.”
What are the consequences of loss of sex-specific territory?
Years ago a man who had worked in PNG told me that fertility shot up when churches convinced men and women to cohabit, where previously they had separate land and houses.
What effect could reduction of habitat, forced cohabitation, forcibly changed migration routes and wiped out populations have on male/female kangaroo territory and consequently on fertility opportunities?
How do we know that the female bias (recorded by Fletcher) at Tinbinburra, for instance, is not due to that area being female territory?
Female elder kangaroos
Daughters seem to learn from their mothers to look after joeys. Where female kangaroos are early orphaned their parenting skill may increase risks in joey upbringing. The extraordinary rates of joey mortality may have something to do with this. (See Faces in the Mob for a study of success and failure in raising joeys in one mob.)
In conclusion, regarding ACT Kangaroo management:
It seems that ACT Roo Management Policy and Science:
- Fails to monitor family structure (spatial population monitoring)
- Fails to deal with size reduction, fertility increases probably related to culls etc
- Fails to look at behaviour; notably breeding limitations exerted through incest avoidance/dominance and separate male/female territory
- Underestimates immigration (See Zenger et al)
- Fails to use DNA monitoring to help in the above
- Seems excessively presumptive and mechanistic
NOTES
[1] Zenger et al DNA study 2003: (Heredity (2003) 91, 153–162. doi:10.1038/sj.hdy.6800293, K R Zenger, M D B Eldridge and D W Cooper, "Intraspecific variation, sex-biased dispersal and phylogeography of the eastern grey kangaroo (Macropus giganteus)."
http://www.nature.com/hdy/journal/v91/n2/full/6800293a.html)
[2] Harvesting impact literature: Many of these studies arise from fish stocks. Articles quoting studies for kangaroos tend to quote from the same very small amount of literature and to draw equivocal conclusions, frequently paraphrasing each other. Peter T. Hale, "Genetic effects of kangaroo harvesting", Australian Mammalogy 26:75-86 (2004)http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.319.7936&rep=rep1&type=pdf seems to be the main work cited, but relies on studies which Fletcher's Phd calls into question, has little to say about eastern grey kangaroos but seems to infer that they have similar rates of starvation attributed to red kangaroos.
In Review of Scientific Literature Relevant to the Commercial Harvest Management of Kangaroos (2011) http://www.environment.nsw.gov.au/resources/nature/110641Kangaroolitreview.pdf , pp.27-28, after flagging the potential impact of harvesting on kangaroos, the study concludes with a mere opinion that the impact of size and other harvesting selection on kangaroos probably would not be great, on the assumption that the harvested populations are not isolated. This is pretty much as Hale's study (above) concludes. However we know that the harvested populations in northern NSW and Queensland are genetically isolated and impoverished according to Zenger et all (2003) cited above. Furthermore, the review showed it was aware of Zenger et al.
"The last two reviews concluded that there was no evidence, or potential, that commercial harvesting could alter the genetic structure of kangaroo populations at current harvesting levels (Olsen and
Braysher 2000, Olsen and Low 2006). It was perceived that kangaroo populations would have to be
reduced to very low levels for genetic impacts to become significant (Olsen and Braysher 2000).
Moreover, at the time of the last review, it was concluded that there was an absence of theoretical,
empirical and modelled evidence of genetic impacts at current levels of harvesting” (Olsen and Low
2006, p50) and there were few, if any,examples of harvest‐induced body size selection in terrestrial
vertebrates. While there have not been any studies specifically investigating the potential genetic
impacts of harvesting kangaroos since the last review, there have been a large number of original
research and review papers addressing this question in a range of other vertebrate species,
highlighting the perception that the potential genetic consequences of harvesting may be significantThe human harvest of wild animals is generally not a random process, with harvesters often
selecting phenotypically desirable animals, e.g. those with a large body size or elaborate weaponry,
such as antlers. This therefore has the potential to impose selective pressure on wild populations,
which may result in an alteration to population structure by reducing the frequency of these
desirable phenotypes and/or an overall loss of genetic variation (Allendorf et al. 2008).
Allendorf and Hard (2009) have termed this process “unnatural selection”, which is defined as undesirable changes in an exploited population due to selection against desirable phenotypes. Cited examples of the
effects of selective harvesting on desirable phenotypes include an increase in the number of tuskless elephants (Loxodonta africana) in South Luanga National Park, Zambia, and a decrease in horn size
of bighorn sheep (Ovis canadensis) because of trophy hunting (reviewed in Allendrof and Hard
2009). In the case of bighorn sheep, the observed genotypic and phenotypic effects resulted from
selective harvesting of young males with rapidly growing horns a trait linked with high reproductive
success) before they reached an age where they could achieve high reproductive success (Coltman
et al. 2003). This study highlights the importance of understanding age-specific trait size, rather
than trait size per se.A recent review by Mysterud (2011) discusses the relative importance of various biotic and abiotic
factors that determine the potential for selective pressure from harvesting. In particular, Mysterud
highlights the importance of assessing selective harvesting within the context of management
regulations, hunting methods, animal trait variance, behaviour and abundance. Mysterud argues that in many cultures large mammal harvesting is not expected to produce strong directional
selection in trait size.Although many of the factors discussed are of greater relevance to traditional
sport hunting, this review highlights the importance of a number of factors relevant to the
commercial harvesting of kangaroos in Australia.There is certainly evidence for selective harvesting of larger/older animals within kangaroo
populations, primarily because the economic performance of kangaroo harvesting enterprises is
highly sensitive to variations in average carcase weight (Stayner 2007). Between 1997 and 2009 the
total harvest in NSW comprised between 70 and 89% males. In the case of wallaroos, the
commercial take is even more strongly biased towards males (almost 90%), because females rarely
reach the minimum size dictated by licence and market conditions (Payne 2011). Despite the
preference for larger males, it was reported that harvesters target a range of sizes above the
minimum, especially when densities are reduced and there are fewer target animals (Payne 2011).
There average weight of harvested animals supports this assertion [(Table 2)].As reported in the last review (Olsen and Low 2006), studies on the potential effects of size-selective harvesting in kangaroos concluded that although there was potential for genetic consequences of
harvesting within a closed population (Tenhumberg et al. 2004), the degree of mobility and
geographic range of genetic populations of kangaroos would be sufficient to ensure that any
localised effects could be countered by immigration (Hale 2004). So, the question remains: does the
recent literature on this topic provide any basis for changing the previous conclusions?Probably not.
In the big horn sheep example referred to above, the extent of selective harvesting pressure was
probably much stronger than occurs in kangaroo populations. In addition, the population was small,
isolated and had restricted potential for immigration (Coltman et al. 2003), thereby exhibiting
characteristics akin to a closed population. As such, this probably represents a more extreme
example, where prevailing management and biological factors combined to create strong selective
pressure."
Penny Olsen and Tim Low, "Situation Analysis Report, Update on Current State of Scientific Knowledge on Kangaroos in the Environment, Including Ecological and Economic Impact and Effect of Culling," School of Botany and Zoology, Australian National University, Canberra, ACT and 6 Henry Street, Chapel Hill, Queensland, Prepared for the Kangaroo Management Advisory Panel, March 2006
Proceedings of the 2010 RSPCA Australia Scientific Seminar: Convergence or conflict: animal welfare in wildlife management and conservation, Tuesday 23 February 2010, CSIRO Discovery Centre, Canberra https://www.rspca.org.au/files/website/The-facts/Science/Scientific-Seminar/2010/SciSem2010-Proceedings.pdf
[3] Family structure/westermarck/incest avoidance/endogamy/exogamy: Sheila Newman, Demography, Territory, Law: Rules of animal and human populations, Countershock Press, 2013, Chapter 3, “CHAPTER 3: The urge to disperse: Why children don’t usually marry their parents.” (Available amazon.com) Examples of incest avoidance citations within:
"Several studies have shown that maternal relatives avoid mating with one another (rhesus macaques: Smith, 1995; red colobus, Procolobus badius temminckii: Starin, 2001; Japanese macaques: Takahata et al., 2002; and see for review: Moore, 1993; van Noordwijk and van Schaik, 2004), 88 but less is known concerning patterns of inbreeding avoidance between paternal relatives (but see Alberts, 1999). In this study, we showed that the probability of paternity by a dominant male decreased when he was related to the dam at R = .5 (the highest possible relatedness coefficient in our study). Smith (1995) showed in rhesus macaques that the intensity of inbreeding avoidance was directly correlated with the closeness of kinship, as in the mandrills studied here. ”
Marie Charpentier, Patricia Peignot, Martine Hossaert-McKey, Olivier Gimenez, Joanna M. Setchell, and E. Jean Wickings., 2005. “Constraints on control: factors influencing reproductive success in male mandrills (Mandrillus sphinx).” Behavioral Ecology 16:614–623]
More reference examples on incest avoidance in multiple species:
Hoier, S., 2003. “Father absence and the age of menarch, A test of four evolutionary models,” Human Nature, Vol. 14, No. 3, pp. 209–233, Walter de Gruyter, Inc., New York.
Cockburn A, Osmond HL, Mulder RA, Green DJ, Douvle MC, 2003. Divorce, dispersal and incest avoidance in the cooperatively breeding superb fairy-wren Malurus cyaneus. J Anim Ecol 185 72:189–202;
Griffin AS, Pemberton JM, Brotherton PNM, McIlrath G, Gaynor D, Kansky R, O'Riain J, Clutton-Brock TH, 2003. A genetic analysis of breeding success in the cooperative meerkat (Suricata suricatta). Behav Ecol 14:472–480;
Mateo JM, 2003. Kin recognition in ground squirrels and other rodents. J Mammal 84:1163–1181;
Pusey A, Wolf M, 1996. Inbreeding avoidance in animals. Trends Ecol Evol 11:201–206;
Stow AJ, Sunnucks P, 2004. Inbreeding avoidance in Cunningham's skinks (Egernia cunninghami) in natural and fragmented habitat. Mol Ecol 13:443–447;
Yu XD, Sun RY, Fang JM, 2004. Effect of kinship on social behaviors in Brandt's voles (Microtus brandti). J Ethol 22:17–22.
Recent comments